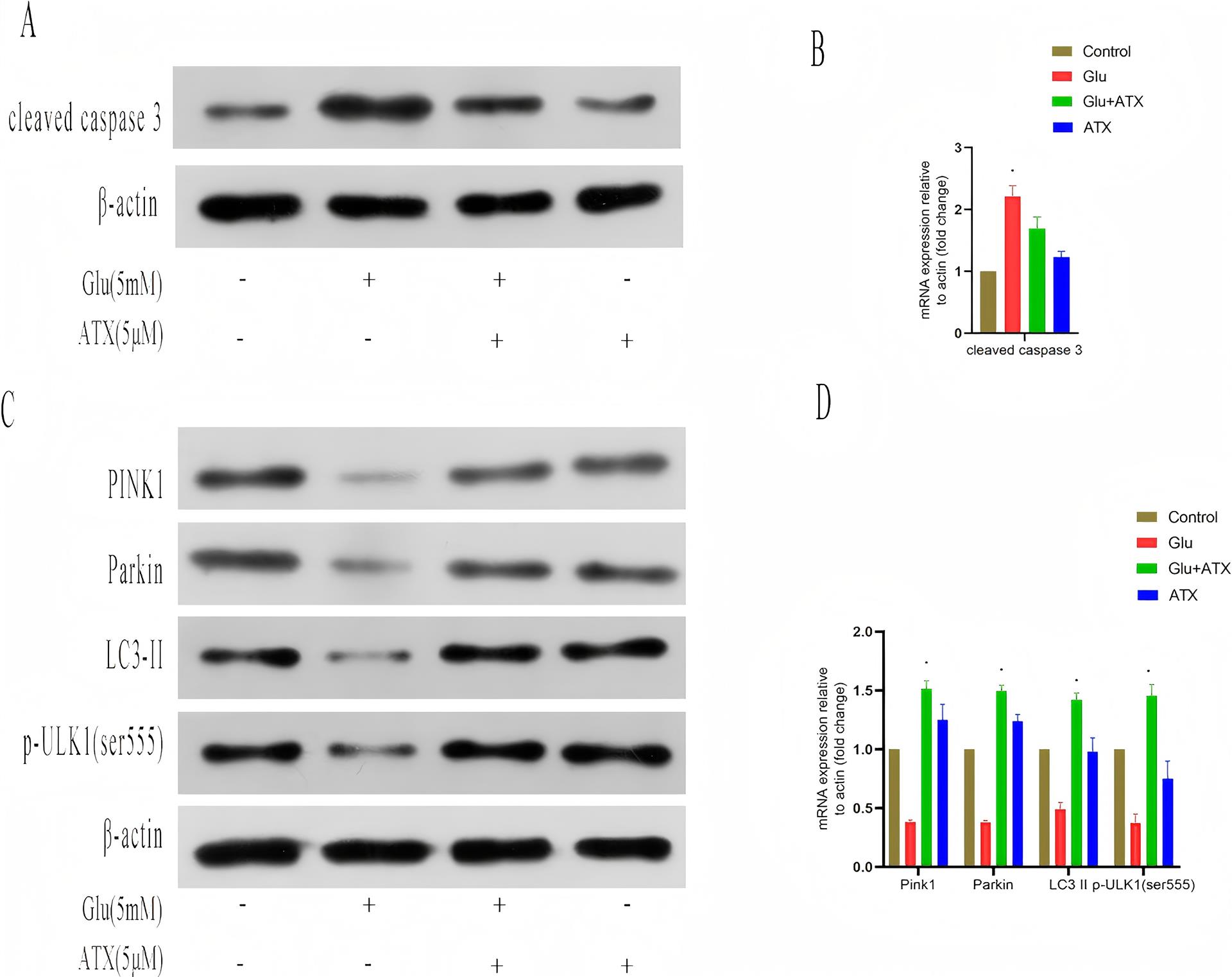
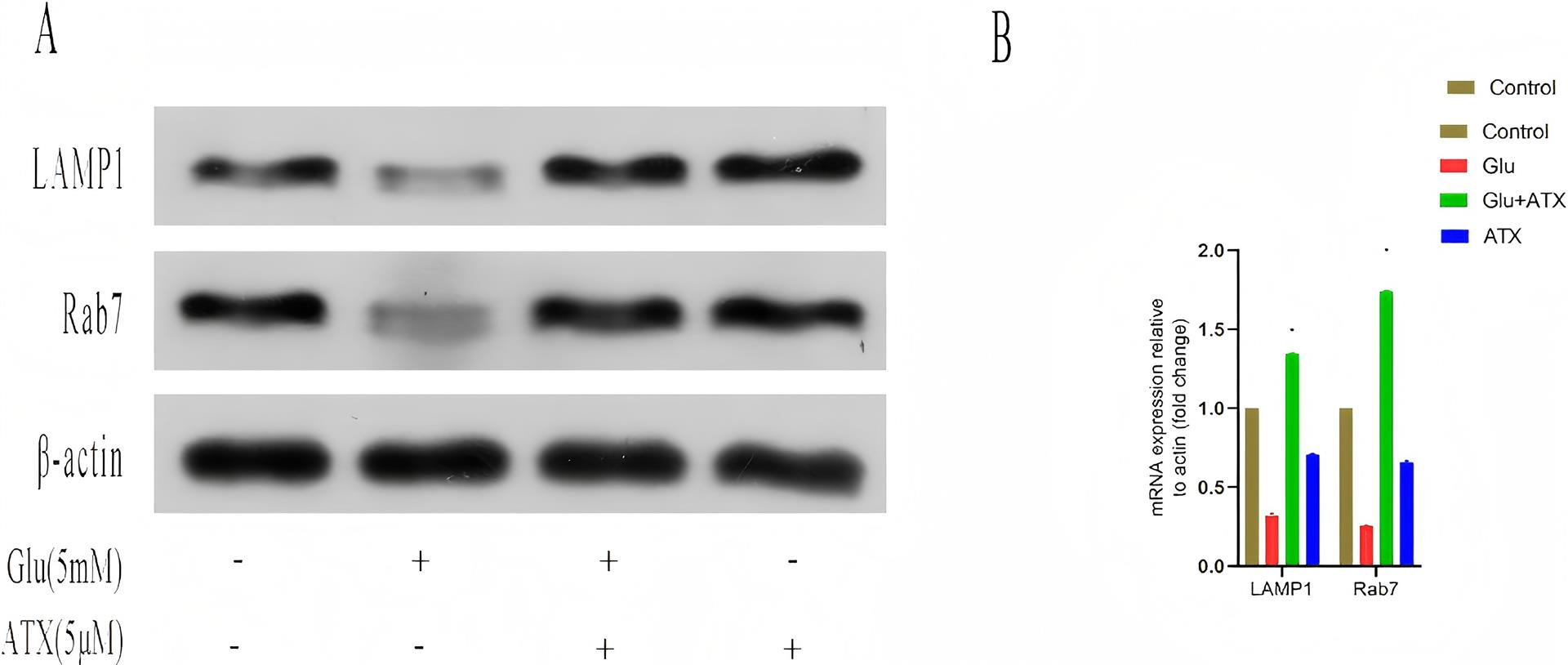
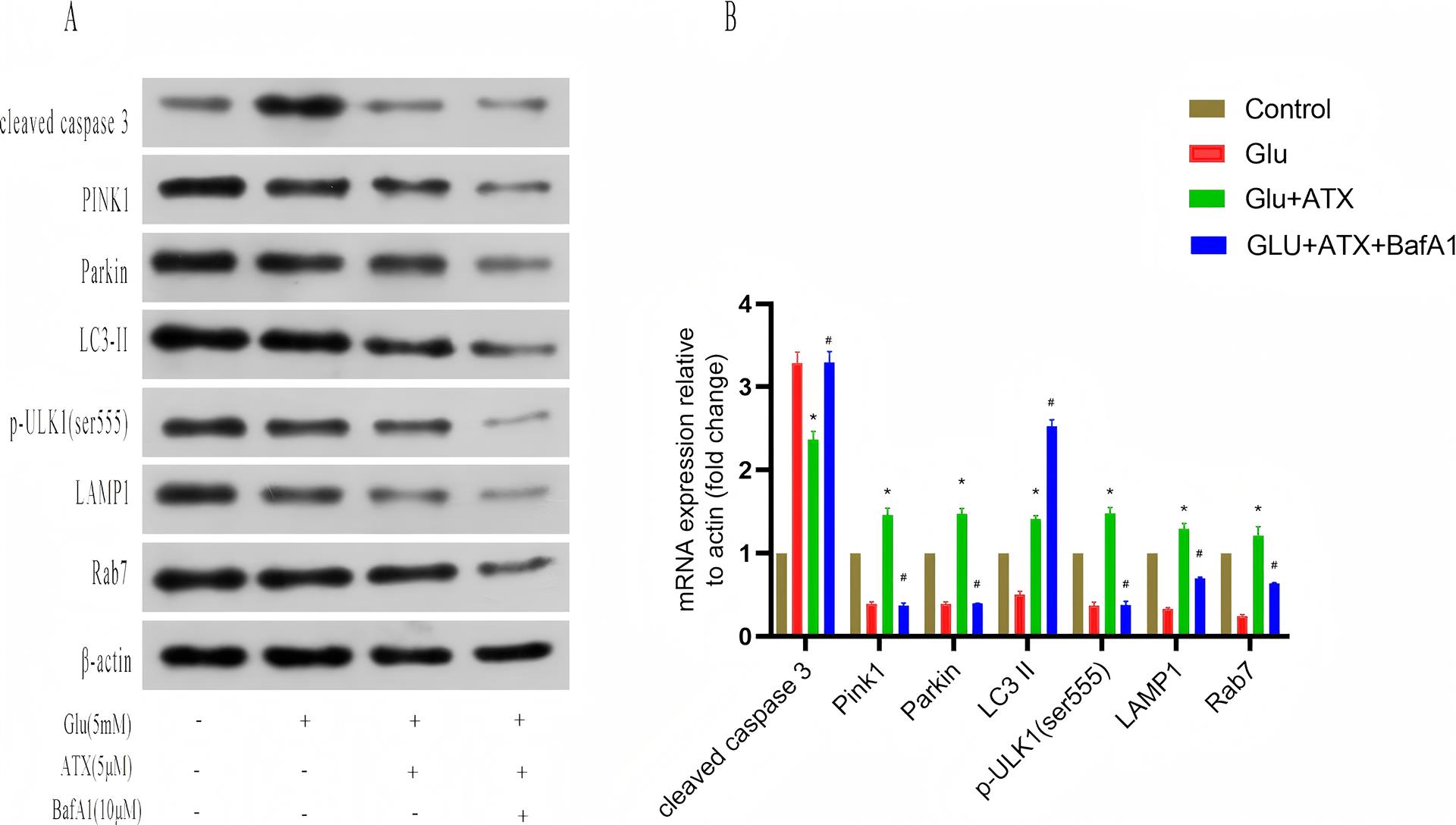
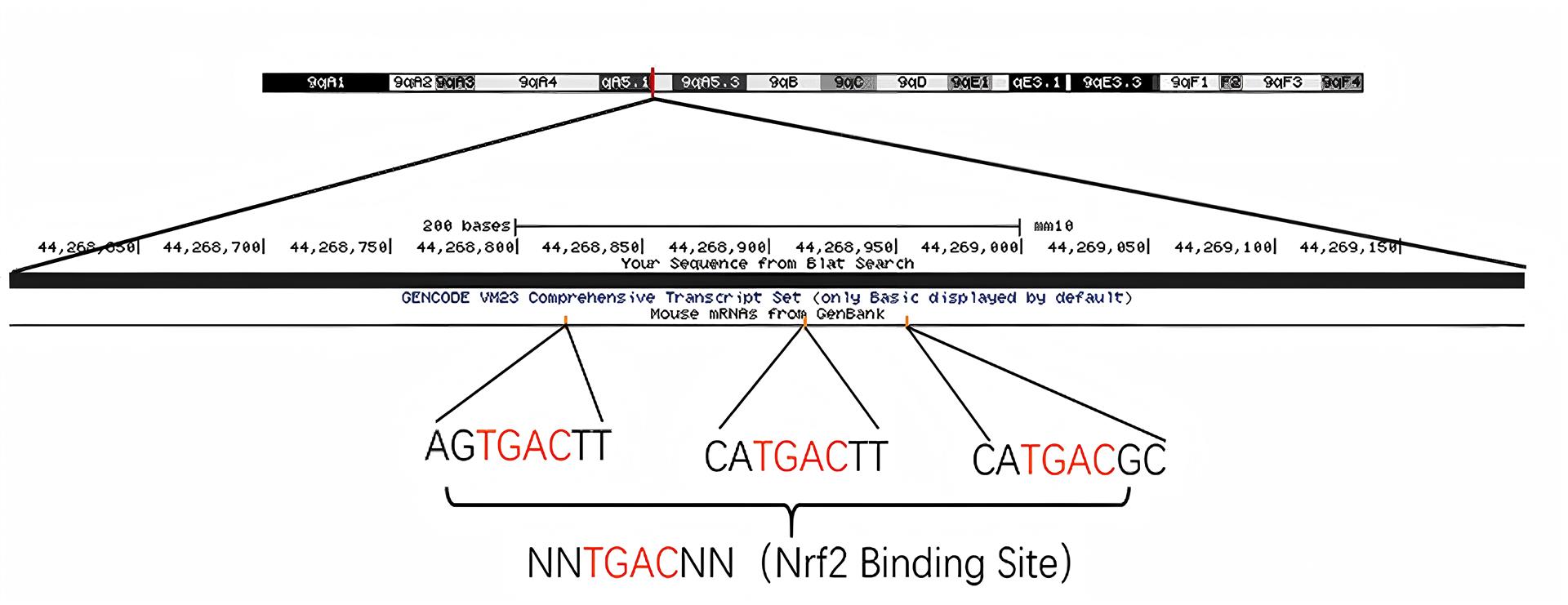
1、JIA%E2%80%83L%EF%BC%8CQUAN%E2%80%83M%EF%BC%8CFU%E2%80%83Y%EF%BC%8Cet%E2%80%83al%EF%BC%8EDementia%E2%80%83in%E2%80%83China%EF%BC%9AEpidemiology%EF%BC%8Cclinical%E2%80%83management%EF%BC%8Cand%E2%80%83%20research%E2%80%83%0Aadvances%EF%BC%BBJ%EF%BC%BD%EF%BC%8ELancet%E2%80%83Neurol%EF%BC%8C2020%EF%BC%8C19%EF%BC%881%EF%BC%89%EF%BC%9A%0A81-92%EF%BC%8EJIA%E2%80%83L%EF%BC%8CQUAN%E2%80%83M%EF%BC%8CFU%E2%80%83Y%EF%BC%8Cet%E2%80%83al%EF%BC%8EDementia%E2%80%83in%E2%80%83China%EF%BC%9AEpidemiology%EF%BC%8Cclinical%E2%80%83management%EF%BC%8Cand%E2%80%83%20research%E2%80%83%0Aadvances%EF%BC%BBJ%EF%BC%BD%EF%BC%8ELancet%E2%80%83Neurol%EF%BC%8C2020%EF%BC%8C19%EF%BC%881%EF%BC%89%EF%BC%9A%0A81-92%EF%BC%8E
2、%E5%91%A8%E5%AE%87%EF%BC%8C%E5%BC%A0%E7%9B%BC%E7%9B%BC%EF%BC%8C%E8%92%8B%E6%80%A1%E5%8D%8E%EF%BC%8C%E7%AD%89%EF%BC%8E%E7%A4%BE%E5%8C%BA%E8%80%81%E4%BA%BA%E8%BD%BB%E5%BA%A6%E8%AE%A4%E7%9F%A5%E5%8A%9F%0A%E8%83%BD%E9%9A%9C%E7%A2%8D%E7%8E%B0%E7%8A%B6%E5%8F%8A%E5%85%B6%E5%BD%B1%E5%93%8D%E5%9B%A0%E7%B4%A0%E9%97%B4%E4%BA%A4%E4%BA%92%E4%BD%9C%E7%94%A8%E5%88%86%E6%9E%90%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0A%E5%B9%BF%E5%B7%9E%E5%8C%BB%E8%8D%AF%EF%BC%8C%E2%80%832023%EF%BC%8C54%EF%BC%888%EF%BC%89%EF%BC%9A52-57%EF%BC%8E%E5%91%A8%E5%AE%87%EF%BC%8C%E5%BC%A0%E7%9B%BC%E7%9B%BC%EF%BC%8C%E8%92%8B%E6%80%A1%E5%8D%8E%EF%BC%8C%E7%AD%89%EF%BC%8E%E7%A4%BE%E5%8C%BA%E8%80%81%E4%BA%BA%E8%BD%BB%E5%BA%A6%E8%AE%A4%E7%9F%A5%E5%8A%9F%0A%E8%83%BD%E9%9A%9C%E7%A2%8D%E7%8E%B0%E7%8A%B6%E5%8F%8A%E5%85%B6%E5%BD%B1%E5%93%8D%E5%9B%A0%E7%B4%A0%E9%97%B4%E4%BA%A4%E4%BA%92%E4%BD%9C%E7%94%A8%E5%88%86%E6%9E%90%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0A%E5%B9%BF%E5%B7%9E%E5%8C%BB%E8%8D%AF%EF%BC%8C%E2%80%832023%EF%BC%8C54%EF%BC%888%EF%BC%89%EF%BC%9A52-57%EF%BC%8E
3、袁行勇,姚春,陈炜,等.阿尔茨海默病不同治疗方式研究新进展[J].广州医药,2024,55(3):236-244.袁行勇,姚春,陈炜,等.阿尔茨海默病不同治疗方式研究新进展[J].广州医药,2024,55(3):236-244.
4、GU%E2%80%83Y%20Y%EF%BC%8CZHAO%E2%80%83X%20R%EF%BC%8CZHANG%E2%80%83N%EF%BC%8Cet%E2%80%83al%EF%BC%8EMitochondrial%E2%80%83%0Adysfunction%E2%80%83as%E2%80%83a%E2%80%83therapeutic%E2%80%83strategy%E2%80%83for%E2%80%83neurodegenerative%E2%80%83%0Adiseases%EF%BC%9ACurrent%E2%80%83insights%E2%80%83and%E2%80%83future%E2%80%83directions%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AAgeing%E2%80%83Res%E2%80%83Rev%EF%BC%8C%E2%80%832024%EF%BC%88102%EF%BC%89%EF%BC%9A102577%EF%BC%8EGU%E2%80%83Y%20Y%EF%BC%8CZHAO%E2%80%83X%20R%EF%BC%8CZHANG%E2%80%83N%EF%BC%8Cet%E2%80%83al%EF%BC%8EMitochondrial%E2%80%83%0Adysfunction%E2%80%83as%E2%80%83a%E2%80%83therapeutic%E2%80%83strategy%E2%80%83for%E2%80%83neurodegenerative%E2%80%83%0Adiseases%EF%BC%9ACurrent%E2%80%83insights%E2%80%83and%E2%80%83future%E2%80%83directions%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AAgeing%E2%80%83Res%E2%80%83Rev%EF%BC%8C%E2%80%832024%EF%BC%88102%EF%BC%89%EF%BC%9A102577%EF%BC%8E
5、TAO%E2%80%83G%EF%BC%8CWANG%E2%80%83X%EF%BC%8CWANG%E2%80%83J%EF%BC%8Cet%E2%80%83al%EF%BC%8EDihydro%02resveratrol%E2%80%83ameliorates%E2%80%83NLRP3%E2%80%83inflammasome-mediated%E2%80%83%0Aneuroinflammation%E2%80%83via%E2%80%83Bnip3-dependent%E2%80%83mitophagy%E2%80%83in%E2%80%83%0AAlzheimer%E2%80%99s%E2%80%83disease%EF%BC%BBJ%EF%BC%BD%EF%BC%8EBr%E2%80%83J%E2%80%83Pharmacol%EF%BC%8C2025%EF%BC%8C%0A182%EF%BC%884%EF%BC%89%EF%BC%9A1005-1024%EF%BC%8ETAO%E2%80%83G%EF%BC%8CWANG%E2%80%83X%EF%BC%8CWANG%E2%80%83J%EF%BC%8Cet%E2%80%83al%EF%BC%8EDihydro%02resveratrol%E2%80%83ameliorates%E2%80%83NLRP3%E2%80%83inflammasome-mediated%E2%80%83%0Aneuroinflammation%E2%80%83via%E2%80%83Bnip3-dependent%E2%80%83mitophagy%E2%80%83in%E2%80%83%0AAlzheimer%E2%80%99s%E2%80%83disease%EF%BC%BBJ%EF%BC%BD%EF%BC%8EBr%E2%80%83J%E2%80%83Pharmacol%EF%BC%8C2025%EF%BC%8C%0A182%EF%BC%884%EF%BC%89%EF%BC%9A1005-1024%EF%BC%8E
6、KIM%E2%80%83S%E2%80%83H%EF%BC%8CKIM%E2%80%83H%EF%BC%8EAstaxanthin%E2%80%83%20modulation%E2%80%83%20of%E2%80%83%0Asignaling%E2%80%83pathways%E2%80%83that%E2%80%83regulate%E2%80%83autophagy%EF%BC%BBJ%EF%BC%BD%EF%BC%8EMar%E2%80%83%0ADrugs%EF%BC%8C2019%EF%BC%8C17%EF%BC%8810%EF%BC%89%EF%BC%9A546%EF%BC%8EKIM%E2%80%83S%E2%80%83H%EF%BC%8CKIM%E2%80%83H%EF%BC%8EAstaxanthin%E2%80%83%20modulation%E2%80%83%20of%E2%80%83%0Asignaling%E2%80%83pathways%E2%80%83that%E2%80%83regulate%E2%80%83autophagy%EF%BC%BBJ%EF%BC%BD%EF%BC%8EMar%E2%80%83%0ADrugs%EF%BC%8C2019%EF%BC%8C17%EF%BC%8810%EF%BC%89%EF%BC%9A546%EF%BC%8E
7、FAKHRI%E2%80%83S%EF%BC%8CANEVA%E2%80%83I%E2%80%83Y%EF%BC%8CFARZAEI%E2%80%83M%E2%80%83H%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AThe%E2%80%83neuroprotective%E2%80%83effects%E2%80%83of%E2%80%83astaxanthin%EF%BC%9ATherapeutic%E2%80%83%0Atargets%E2%80%83and%E2%80%83clinical%E2%80%83perspective%EF%BC%BBJ%EF%BC%BD%EF%BC%8EMolecules%EF%BC%8C%0A2019%EF%BC%8C24%EF%BC%8814%EF%BC%89%EF%BC%9A2640%EF%BC%8EFAKHRI%E2%80%83S%EF%BC%8CANEVA%E2%80%83I%E2%80%83Y%EF%BC%8CFARZAEI%E2%80%83M%E2%80%83H%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AThe%E2%80%83neuroprotective%E2%80%83effects%E2%80%83of%E2%80%83astaxanthin%EF%BC%9ATherapeutic%E2%80%83%0Atargets%E2%80%83and%E2%80%83clinical%E2%80%83perspective%EF%BC%BBJ%EF%BC%BD%EF%BC%8EMolecules%EF%BC%8C%0A2019%EF%BC%8C24%EF%BC%8814%EF%BC%89%EF%BC%9A2640%EF%BC%8E
8、ZHANG%E2%80%83Y%EF%BC%8CYAO%E2%80%83Y%EF%BC%8CQIU%E2%80%83X%EF%BC%8Cet%E2%80%83al%EF%BC%8EListeria%E2%80%83hijacks%E2%80%83%0Ahost%E2%80%83mitophagy%E2%80%83through%E2%80%83%20a%E2%80%83%20novel%E2%80%83mitophagy%E2%80%83%20receptor%E2%80%83%0Ato%E2%80%83evade%E2%80%83killing%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENat%E2%80%83Immunol%EF%BC%8C%E2%80%832019%EF%BC%8C20%0A%EF%BC%884%EF%BC%89%EF%BC%9A433-446%EF%BC%8EZHANG%E2%80%83Y%EF%BC%8CYAO%E2%80%83Y%EF%BC%8CQIU%E2%80%83X%EF%BC%8Cet%E2%80%83al%EF%BC%8EListeria%E2%80%83hijacks%E2%80%83%0Ahost%E2%80%83mitophagy%E2%80%83through%E2%80%83%20a%E2%80%83%20novel%E2%80%83mitophagy%E2%80%83%20receptor%E2%80%83%0Ato%E2%80%83evade%E2%80%83killing%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENat%E2%80%83Immunol%EF%BC%8C%E2%80%832019%EF%BC%8C20%0A%EF%BC%884%EF%BC%89%EF%BC%9A433-446%EF%BC%8E
9、MOORE%E2%80%83CB%EF%BC%8CBERGSTRALH%E2%80%83DT%EF%BC%8CDUNCAN%E2%80%83JA%EF%BC%8C%0Aet%E2%80%83al%EF%BC%8ENLRX1%E2%80%83is%E2%80%83a%E2%80%83%20regulator%E2%80%83of%E2%80%83mitochondrial%E2%80%83antiviral%E2%80%83%0Aimmunity%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENature%EF%BC%8C2008%EF%BC%8C451%EF%BC%887178%EF%BC%89%EF%BC%9A%0A573-577%EF%BC%8EMOORE%E2%80%83CB%EF%BC%8CBERGSTRALH%E2%80%83DT%EF%BC%8CDUNCAN%E2%80%83JA%EF%BC%8C%0Aet%E2%80%83al%EF%BC%8ENLRX1%E2%80%83is%E2%80%83a%E2%80%83%20regulator%E2%80%83of%E2%80%83mitochondrial%E2%80%83antiviral%E2%80%83%0Aimmunity%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENature%EF%BC%8C2008%EF%BC%8C451%EF%BC%887178%EF%BC%89%EF%BC%9A%0A573-577%EF%BC%8E
10、STOKMAN%E2%80%83G%EF%BC%8CKORS%E2%80%83L%EF%BC%8CBAKKER%E2%80%83PJ%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ANLRX1%E2%80%83%20dampens%E2%80%83%20oxidative%E2%80%83%20stress%E2%80%83%20and%E2%80%83%20apoptosis%E2%80%83in%E2%80%83%0Atissue%E2%80%83injury%E2%80%83via%E2%80%83control%E2%80%83of%E2%80%83mitochondrial%E2%80%83activity%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AJ%E2%80%83Exp%E2%80%83Med%EF%BC%8C2017%EF%BC%8C214%EF%BC%888%EF%BC%89%EF%BC%9A2405-2420%EF%BC%8ESTOKMAN%E2%80%83G%EF%BC%8CKORS%E2%80%83L%EF%BC%8CBAKKER%E2%80%83PJ%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ANLRX1%E2%80%83%20dampens%E2%80%83%20oxidative%E2%80%83%20stress%E2%80%83%20and%E2%80%83%20apoptosis%E2%80%83in%E2%80%83%0Atissue%E2%80%83injury%E2%80%83via%E2%80%83control%E2%80%83of%E2%80%83mitochondrial%E2%80%83activity%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AJ%E2%80%83Exp%E2%80%83Med%EF%BC%8C2017%EF%BC%8C214%EF%BC%888%EF%BC%89%EF%BC%9A2405-2420%EF%BC%8E
11、NAGAI-SINGER%E2%80%83M%E2%80%83A%EF%BC%8CMORRISON%E2%80%83H%E2%80%83A%EF%BC%8CALLEN%E2%80%83%20I%E2%80%83%0AC%EF%BC%8ENLRX1%E2%80%83is%E2%80%83a%E2%80%83multifaceted%E2%80%83and%E2%80%83enigmatic%E2%80%83regulator%E2%80%83of%E2%80%83%0Aimmune%E2%80%83system%E2%80%83function%EF%BC%BBJ%EF%BC%BD%EF%BC%8EFront%E2%80%83Immunol%EF%BC%8C2019%0A%EF%BC%8810%EF%BC%89%EF%BC%9A2419%EF%BC%8ENAGAI-SINGER%E2%80%83M%E2%80%83A%EF%BC%8CMORRISON%E2%80%83H%E2%80%83A%EF%BC%8CALLEN%E2%80%83%20I%E2%80%83%0AC%EF%BC%8ENLRX1%E2%80%83is%E2%80%83a%E2%80%83multifaceted%E2%80%83and%E2%80%83enigmatic%E2%80%83regulator%E2%80%83of%E2%80%83%0Aimmune%E2%80%83system%E2%80%83function%EF%BC%BBJ%EF%BC%BD%EF%BC%8EFront%E2%80%83Immunol%EF%BC%8C2019%0A%EF%BC%8810%EF%BC%89%EF%BC%9A2419%EF%BC%8E
12、GILL%E2%80%83A%20J%EF%BC%8CSMITH%E2%80%83M%20D%EF%BC%8CGALLEGUILLOS%E2%80%83D%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ANLRX1%E2%80%83limits%E2%80%83inflammatory%E2%80%83neurodegeneration%E2%80%83in%E2%80%83the%E2%80%83%0Aanterior%E2%80%83visual%E2%80%83pathway%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Neuroinflammation%EF%BC%8C%0A2025%EF%BC%8C22%EF%BC%881%EF%BC%89%EF%BC%9A21%EF%BC%8EGILL%E2%80%83A%20J%EF%BC%8CSMITH%E2%80%83M%20D%EF%BC%8CGALLEGUILLOS%E2%80%83D%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ANLRX1%E2%80%83limits%E2%80%83inflammatory%E2%80%83neurodegeneration%E2%80%83in%E2%80%83the%E2%80%83%0Aanterior%E2%80%83visual%E2%80%83pathway%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Neuroinflammation%EF%BC%8C%0A2025%EF%BC%8C22%EF%BC%881%EF%BC%89%EF%BC%9A21%EF%BC%8E
13、%E2%80%83%20PAJARES%E2%80%83M%EF%BC%8CJIM%C3%89NEZ-MORENO%E2%80%83N%EF%BC%8CGARC%C3%8DA%02YAG%C3%9CE%E2%80%83%C3%81%E2%80%83%20J%EF%BC%8Cet%E2%80%83al%EF%BC%8ETranscription%E2%80%83factor%E2%80%83NFE2L2%2F%0ANRF2%E2%80%83is%E2%80%83a%E2%80%83regulator%E2%80%83of%E2%80%83macroautophagy%E2%80%83genes%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AAutophagy%EF%BC%8C2016%EF%BC%8C12%EF%BC%8810%EF%BC%89%EF%BC%9A1902-1916%EF%BC%8E%E2%80%83%20PAJARES%E2%80%83M%EF%BC%8CJIM%C3%89NEZ-MORENO%E2%80%83N%EF%BC%8CGARC%C3%8DA%02YAG%C3%9CE%E2%80%83%C3%81%E2%80%83%20J%EF%BC%8Cet%E2%80%83al%EF%BC%8ETranscription%E2%80%83factor%E2%80%83NFE2L2%2F%0ANRF2%E2%80%83is%E2%80%83a%E2%80%83regulator%E2%80%83of%E2%80%83macroautophagy%E2%80%83genes%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AAutophagy%EF%BC%8C2016%EF%BC%8C12%EF%BC%8810%EF%BC%89%EF%BC%9A1902-1916%EF%BC%8E
14、WEN%E2%80%83X%EF%BC%8CHUANG%E2%80%83A%EF%BC%8CHU%E2%80%83J%EF%BC%8Cet%E2%80%83al%EF%BC%8ENeuroprotective%E2%80%83%0Aeffect%E2%80%83%20of%E2%80%83%20astaxanthin%E2%80%83%20against%E2%80%83%20glutamate-induced%E2%80%83%0Acytotoxicity%E2%80%83in%E2%80%83HT22%E2%80%83cells%EF%BC%9AInvolvement%E2%80%83of%E2%80%83the%E2%80%83Akt%2F%0AGSK-3%CE%B2%E2%80%83pathway%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENeuroscience%EF%BC%8C2015%0A%EF%BC%88303%EF%BC%89%EF%BC%9A558-568%EF%BC%8EWEN%E2%80%83X%EF%BC%8CHUANG%E2%80%83A%EF%BC%8CHU%E2%80%83J%EF%BC%8Cet%E2%80%83al%EF%BC%8ENeuroprotective%E2%80%83%0Aeffect%E2%80%83%20of%E2%80%83%20astaxanthin%E2%80%83%20against%E2%80%83%20glutamate-induced%E2%80%83%0Acytotoxicity%E2%80%83in%E2%80%83HT22%E2%80%83cells%EF%BC%9AInvolvement%E2%80%83of%E2%80%83the%E2%80%83Akt%2F%0AGSK-3%CE%B2%E2%80%83pathway%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENeuroscience%EF%BC%8C2015%0A%EF%BC%88303%EF%BC%89%EF%BC%9A558-568%EF%BC%8E
15、RAMSEY%E2%80%83C%E2%80%83P%EF%BC%8CGLASS%E2%80%83C%E2%80%83A%EF%BC%8CMONTGOMERY%E2%80%83M%E2%80%83B%EF%BC%8C%0Aet%E2%80%83al%EF%BC%8EExpression%E2%80%83of%E2%80%83Nrf2%E2%80%83in%E2%80%83neurodegenerative%E2%80%83diseases%0A%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Neuropathol%E2%80%83Exp%E2%80%83Neurol%EF%BC%8C2007%EF%BC%8C66%EF%BC%881%EF%BC%89%EF%BC%9A%0A75-85%EF%BC%8ERAMSEY%E2%80%83C%E2%80%83P%EF%BC%8CGLASS%E2%80%83C%E2%80%83A%EF%BC%8CMONTGOMERY%E2%80%83M%E2%80%83B%EF%BC%8C%0Aet%E2%80%83al%EF%BC%8EExpression%E2%80%83of%E2%80%83Nrf2%E2%80%83in%E2%80%83neurodegenerative%E2%80%83diseases%0A%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Neuropathol%E2%80%83Exp%E2%80%83Neurol%EF%BC%8C2007%EF%BC%8C66%EF%BC%881%EF%BC%89%EF%BC%9A%0A75-85%EF%BC%8E
16、BAHN%E2%80%83G%EF%BC%8CJO%E2%80%83D%E2%80%83G%EF%BC%8ETherapeutic%E2%80%83%20approaches%E2%80%83%20to%E2%80%83%0AAlzheimer%E2%80%99s%E2%80%83disease%E2%80%83through%E2%80%83modulation%E2%80%83of%E2%80%83NRF2%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ANeuromolecular%E2%80%83Med%EF%BC%8C2019%EF%BC%8C21%EF%BC%881%EF%BC%89%EF%BC%9A1-11%EF%BC%8EBAHN%E2%80%83G%EF%BC%8CJO%E2%80%83D%E2%80%83G%EF%BC%8ETherapeutic%E2%80%83%20approaches%E2%80%83%20to%E2%80%83%0AAlzheimer%E2%80%99s%E2%80%83disease%E2%80%83through%E2%80%83modulation%E2%80%83of%E2%80%83NRF2%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ANeuromolecular%E2%80%83Med%EF%BC%8C2019%EF%BC%8C21%EF%BC%881%EF%BC%89%EF%BC%9A1-11%EF%BC%8E
17、XU%E2%80%83J%EF%BC%8CZHOU%E2%80%83L%EF%BC%8CWENG%E2%80%83Q%EF%BC%8Cet%E2%80%83al%EF%BC%8ECu%20rcumin%E2%80%83%0Aanalogues%E2%80%83attenuate%E2%80%83A%CE%B225-35-induced%E2%80%83%20oxidative%E2%80%83%0Astress%E2%80%83in%E2%80%83PC12%E2%80%83cells%E2%80%83via%E2%80%83Keap1%2FNrf2%2FHO-1%E2%80%83%20signaling%E2%80%83%0Apathways%EF%BC%BBJ%EF%BC%BD%EF%BC%8EChem%E2%80%83Biol%E2%80%83Interact%EF%BC%8C2019%EF%BC%88305%EF%BC%89%EF%BC%9A%0A171-179%EF%BC%8EXU%E2%80%83J%EF%BC%8CZHOU%E2%80%83L%EF%BC%8CWENG%E2%80%83Q%EF%BC%8Cet%E2%80%83al%EF%BC%8ECu%20rcumin%E2%80%83%0Aanalogues%E2%80%83attenuate%E2%80%83A%CE%B225-35-induced%E2%80%83%20oxidative%E2%80%83%0Astress%E2%80%83in%E2%80%83PC12%E2%80%83cells%E2%80%83via%E2%80%83Keap1%2FNrf2%2FHO-1%E2%80%83%20signaling%E2%80%83%0Apathways%EF%BC%BBJ%EF%BC%BD%EF%BC%8EChem%E2%80%83Biol%E2%80%83Interact%EF%BC%8C2019%EF%BC%88305%EF%BC%89%EF%BC%9A%0A171-179%EF%BC%8E
18、%E2%80%83%20CUI%E2%80%83X%EF%BC%8CZONG%E2%80%83S%EF%BC%8CSONG%E2%80%83W%EF%BC%8Cet%E2%80%83al%EF%BC%8EOmaveloxolone%E2%80%83%0Aameliorates%E2%80%83cognitive%E2%80%83dysfunction%E2%80%83in%E2%80%83APP%2FPS1%E2%80%83mice%E2%80%83by%E2%80%83%0Astabilizing%E2%80%83the%E2%80%83STAT3%E2%80%83pathway%EF%BC%BBJ%EF%BC%BD%EF%BC%8ELife%E2%80%83Sci%EF%BC%8C2023%0A%EF%BC%88335%EF%BC%89%EF%BC%9A122261%EF%BC%8E%E2%80%83%20CUI%E2%80%83X%EF%BC%8CZONG%E2%80%83S%EF%BC%8CSONG%E2%80%83W%EF%BC%8Cet%E2%80%83al%EF%BC%8EOmaveloxolone%E2%80%83%0Aameliorates%E2%80%83cognitive%E2%80%83dysfunction%E2%80%83in%E2%80%83APP%2FPS1%E2%80%83mice%E2%80%83by%E2%80%83%0Astabilizing%E2%80%83the%E2%80%83STAT3%E2%80%83pathway%EF%BC%BBJ%EF%BC%BD%EF%BC%8ELife%E2%80%83Sci%EF%BC%8C2023%0A%EF%BC%88335%EF%BC%89%EF%BC%9A122261%EF%BC%8E
19、DONG%E2%80%83X%E2%80%83X%EF%BC%8CWANG%E2%80%83Y%EF%BC%8CQIN%E2%80%83Z%E2%80%83H%EF%BC%8EMolecula%20r%E2%80%83%0Amechanisms%E2%80%83%20of%E2%80%83%20excitotoxicity%E2%80%83%20and%E2%80%83their%E2%80%83%20relevance%E2%80%83to%E2%80%83%0Apathogenesis%E2%80%83of%E2%80%83neurodegenerative%E2%80%83diseases%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AActa%E2%80%83Pharmacol%E2%80%83Sin%EF%BC%8C%E2%80%832009%EF%BC%8C30%EF%BC%884%EF%BC%89%EF%BC%9A379-387%EF%BC%8EDONG%E2%80%83X%E2%80%83X%EF%BC%8CWANG%E2%80%83Y%EF%BC%8CQIN%E2%80%83Z%E2%80%83H%EF%BC%8EMolecula%20r%E2%80%83%0Amechanisms%E2%80%83%20of%E2%80%83%20excitotoxicity%E2%80%83%20and%E2%80%83their%E2%80%83%20relevance%E2%80%83to%E2%80%83%0Apathogenesis%E2%80%83of%E2%80%83neurodegenerative%E2%80%83diseases%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AActa%E2%80%83Pharmacol%E2%80%83Sin%EF%BC%8C%E2%80%832009%EF%BC%8C30%EF%BC%884%EF%BC%89%EF%BC%9A379-387%EF%BC%8E
20、FAROOQUI%E2%80%83T%EF%BC%8CFAROOQUI%E2%80%83A%EF%BC%8EAging%EF%BC%9Aan%E2%80%83important%E2%80%83%0Afactor%E2%80%83for%E2%80%83the%E2%80%83pathogenesis%E2%80%83of%E2%80%83neurodegenerative%E2%80%83diseases%0A%EF%BC%BBJ%EF%BC%BD%EF%BC%8EMech%E2%80%83Ageing%E2%80%83Dev%EF%BC%8C2009%EF%BC%8C130%EF%BC%884%EF%BC%89%EF%BC%9A203-215%EF%BC%8EFAROOQUI%E2%80%83T%EF%BC%8CFAROOQUI%E2%80%83A%EF%BC%8EAging%EF%BC%9Aan%E2%80%83important%E2%80%83%0Afactor%E2%80%83for%E2%80%83the%E2%80%83pathogenesis%E2%80%83of%E2%80%83neurodegenerative%E2%80%83diseases%0A%EF%BC%BBJ%EF%BC%BD%EF%BC%8EMech%E2%80%83Ageing%E2%80%83Dev%EF%BC%8C2009%EF%BC%8C130%EF%BC%884%EF%BC%89%EF%BC%9A203-215%EF%BC%8E
21、FANG%E2%80%83E%E2%80%83F%EF%BC%8CHOU%E2%80%83Y%EF%BC%8CPALIKARAS%E2%80%83K%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AMitophagy%E2%80%83inhibits%E2%80%83amyloid-%CE%B2%E2%80%83and%E2%80%83tau%E2%80%83pathology%E2%80%83and%E2%80%83%0Areverses%E2%80%83cognitive%E2%80%83deficits%E2%80%83in%E2%80%83models%E2%80%83of%E2%80%83Alzheimer%E2%80%99s%E2%80%83%0Adisease%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENat%E2%80%83Neurosci%EF%BC%8C2019%EF%BC%8C22%EF%BC%883%EF%BC%89%EF%BC%9A401-%0A412%EF%BC%8EFANG%E2%80%83E%E2%80%83F%EF%BC%8CHOU%E2%80%83Y%EF%BC%8CPALIKARAS%E2%80%83K%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AMitophagy%E2%80%83inhibits%E2%80%83amyloid-%CE%B2%E2%80%83and%E2%80%83tau%E2%80%83pathology%E2%80%83and%E2%80%83%0Areverses%E2%80%83cognitive%E2%80%83deficits%E2%80%83in%E2%80%83models%E2%80%83of%E2%80%83Alzheimer%E2%80%99s%E2%80%83%0Adisease%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENat%E2%80%83Neurosci%EF%BC%8C2019%EF%BC%8C22%EF%BC%883%EF%BC%89%EF%BC%9A401-%0A412%EF%BC%8E
22、XIE%E2%80%83C%EF%BC%8CAMAN%E2%80%83Y%EF%BC%8CADRIAANSE%E2%80%83B%E2%80%83A%EF%BC%8Cet%E2%80%83al%EF%BC%8ECulprit%E2%80%83%0Aor%E2%80%83bystander%EF%BC%9ADefective%E2%80%83mitophagy%E2%80%83in%E2%80%83Alzheimer%E2%80%99s%E2%80%83%0Adisease%EF%BC%BBJ%EF%BC%BD%EF%BC%8EFront%E2%80%83Cell%E2%80%83Dev%E2%80%83Biol%EF%BC%8C2020%EF%BC%887%EF%BC%89%EF%BC%9A391%EF%BC%8EXIE%E2%80%83C%EF%BC%8CAMAN%E2%80%83Y%EF%BC%8CADRIAANSE%E2%80%83B%E2%80%83A%EF%BC%8Cet%E2%80%83al%EF%BC%8ECulprit%E2%80%83%0Aor%E2%80%83bystander%EF%BC%9ADefective%E2%80%83mitophagy%E2%80%83in%E2%80%83Alzheimer%E2%80%99s%E2%80%83%0Adisease%EF%BC%BBJ%EF%BC%BD%EF%BC%8EFront%E2%80%83Cell%E2%80%83Dev%E2%80%83Biol%EF%BC%8C2020%EF%BC%887%EF%BC%89%EF%BC%9A391%EF%BC%8E
23、YE%E2%80%83X%EF%BC%8CSUN%E2%80%83X%EF%BC%8CSTAROVOYTOV%E2%80%83V%EF%BC%8Cet%E2%80%83al%EF%BC%8EParkin%02mediated%E2%80%83%20mitophagy%E2%80%83in%E2%80%83%20mutant%E2%80%83%20hAPP%E2%80%83%20neurons%E2%80%83%20and%E2%80%83%0AAlzheimer%E2%80%99s%E2%80%83disease%E2%80%83patient%E2%80%83brains%EF%BC%BBJ%EF%BC%BD%EF%BC%8EHum%E2%80%83Mol%E2%80%83%0AGenet%EF%BC%8C%E2%80%832015%EF%BC%8C24%EF%BC%8810%EF%BC%89%EF%BC%9A2938-2951%EF%BC%8EYE%E2%80%83X%EF%BC%8CSUN%E2%80%83X%EF%BC%8CSTAROVOYTOV%E2%80%83V%EF%BC%8Cet%E2%80%83al%EF%BC%8EParkin%02mediated%E2%80%83%20mitophagy%E2%80%83in%E2%80%83%20mutant%E2%80%83%20hAPP%E2%80%83%20neurons%E2%80%83%20and%E2%80%83%0AAlzheimer%E2%80%99s%E2%80%83disease%E2%80%83patient%E2%80%83brains%EF%BC%BBJ%EF%BC%BD%EF%BC%8EHum%E2%80%83Mol%E2%80%83%0AGenet%EF%BC%8C%E2%80%832015%EF%BC%8C24%EF%BC%8810%EF%BC%89%EF%BC%9A2938-2951%EF%BC%8E
24、%E2%80%83%20IMBEAULT%E2%80%83E%EF%BC%8CMAHVELATI%E2%80%83TM%EF%BC%8CBRAUN%E2%80%83R%EF%BC%8Cet%E2%80%83%0Aal%EF%BC%8ENlrx1%E2%80%83regulates%E2%80%83neuronal%E2%80%83cell%E2%80%83death%EF%BC%BBJ%EF%BC%BD%EF%BC%8EMol%E2%80%83%0ABrain%EF%BC%8C2014%EF%BC%887%EF%BC%89%EF%BC%9A90%EF%BC%8E%E2%80%83%20IMBEAULT%E2%80%83E%EF%BC%8CMAHVELATI%E2%80%83TM%EF%BC%8CBRAUN%E2%80%83R%EF%BC%8Cet%E2%80%83%0Aal%EF%BC%8ENlrx1%E2%80%83regulates%E2%80%83neuronal%E2%80%83cell%E2%80%83death%EF%BC%BBJ%EF%BC%BD%EF%BC%8EMol%E2%80%83%0ABrain%EF%BC%8C2014%EF%BC%887%EF%BC%89%EF%BC%9A90%EF%BC%8E
25、WANG%E2%80%83Y%EF%BC%8CLONG%E2%80%83L%EF%BC%8CLUO%E2%80%83Q%EF%BC%8Cet%E2%80%83al%EF%BC%8EAflatoxin%E2%80%83B1%E2%80%83%0Ainduces%E2%80%83ROS-dependent%E2%80%83mitophagy%E2%80%83by%E2%80%83modulating%E2%80%83the%E2%80%83%0APINK1%2FParkin%E2%80%83pathway%E2%80%83in%E2%80%83HepG2%E2%80%83cells%EF%BC%BBJ%EF%BC%BD%EF%BC%8EBasic%E2%80%83%0AClin%E2%80%83Pharmacol%E2%80%83Toxicol%EF%BC%8C2024%EF%BC%8C135%EF%BC%882%EF%BC%89%EF%BC%9A195-209%EF%BC%8EWANG%E2%80%83Y%EF%BC%8CLONG%E2%80%83L%EF%BC%8CLUO%E2%80%83Q%EF%BC%8Cet%E2%80%83al%EF%BC%8EAflatoxin%E2%80%83B1%E2%80%83%0Ainduces%E2%80%83ROS-dependent%E2%80%83mitophagy%E2%80%83by%E2%80%83modulating%E2%80%83the%E2%80%83%0APINK1%2FParkin%E2%80%83pathway%E2%80%83in%E2%80%83HepG2%E2%80%83cells%EF%BC%BBJ%EF%BC%BD%EF%BC%8EBasic%E2%80%83%0AClin%E2%80%83Pharmacol%E2%80%83Toxicol%EF%BC%8C2024%EF%BC%8C135%EF%BC%882%EF%BC%89%EF%BC%9A195-209%EF%BC%8E
26、YE%E2%80%83H%EF%BC%8CCAI%E2%80%83T%EF%BC%8CSHEN%E2%80%83Y%EF%BC%8Cet%E2%80%83al%EF%BC%8EMST1%E2%80%83%20knockdown%E2%80%83%0Ainhibits%E2%80%83osteoarthritis%E2%80%83progression%E2%80%83through%E2%80%83Parkin%02mediated%E2%80%83mitophagy%E2%80%83and%E2%80%83Nrf2%2FNF-%CE%BAB%E2%80%83%20signalling%E2%80%83%0Apathway%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Cell%E2%80%83Mol%E2%80%83Med%EF%BC%8C2024%EF%BC%8C28%EF%BC%8811%EF%BC%89%EF%BC%9A%0Ae18476%EF%BC%8EYE%E2%80%83H%EF%BC%8CCAI%E2%80%83T%EF%BC%8CSHEN%E2%80%83Y%EF%BC%8Cet%E2%80%83al%EF%BC%8EMST1%E2%80%83%20knockdown%E2%80%83%0Ainhibits%E2%80%83osteoarthritis%E2%80%83progression%E2%80%83through%E2%80%83Parkin%02mediated%E2%80%83mitophagy%E2%80%83and%E2%80%83Nrf2%2FNF-%CE%BAB%E2%80%83%20signalling%E2%80%83%0Apathway%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Cell%E2%80%83Mol%E2%80%83Med%EF%BC%8C2024%EF%BC%8C28%EF%BC%8811%EF%BC%89%EF%BC%9A%0Ae18476%EF%BC%8E
27、SONG%E2%80%83Y%EF%BC%8CLIANG%E2%80%83H%EF%BC%8CLI%E2%80%83G%EF%BC%8Cet%E2%80%83al%EF%BC%8EThe%E2%80%83NLRX1-%0ASLC39A7%E2%80%83complex%E2%80%83orchestrates%E2%80%83mitochondrial%E2%80%83dynamics%E2%80%83%0Aand%E2%80%83mitophagy%E2%80%83to%E2%80%83%20rejuvenate%E2%80%83intervertebral%E2%80%83%20disc%E2%80%83%20by%E2%80%83%0Amodulating%E2%80%83mitochondrial%E2%80%83Zn2%2B%E2%80%83trafficking%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AAutophagy%EF%BC%8C2024%EF%BC%8C20%EF%BC%884%EF%BC%89%EF%BC%9A809-829%EF%BC%8ESONG%E2%80%83Y%EF%BC%8CLIANG%E2%80%83H%EF%BC%8CLI%E2%80%83G%EF%BC%8Cet%E2%80%83al%EF%BC%8EThe%E2%80%83NLRX1-%0ASLC39A7%E2%80%83complex%E2%80%83orchestrates%E2%80%83mitochondrial%E2%80%83dynamics%E2%80%83%0Aand%E2%80%83mitophagy%E2%80%83to%E2%80%83%20rejuvenate%E2%80%83intervertebral%E2%80%83%20disc%E2%80%83%20by%E2%80%83%0Amodulating%E2%80%83mitochondrial%E2%80%83Zn2%2B%E2%80%83trafficking%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AAutophagy%EF%BC%8C2024%EF%BC%8C20%EF%BC%884%EF%BC%89%EF%BC%9A809-829%EF%BC%8E