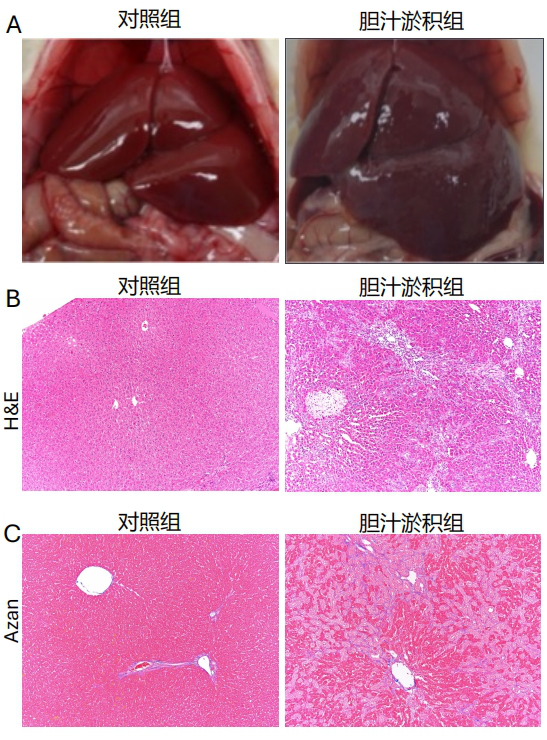
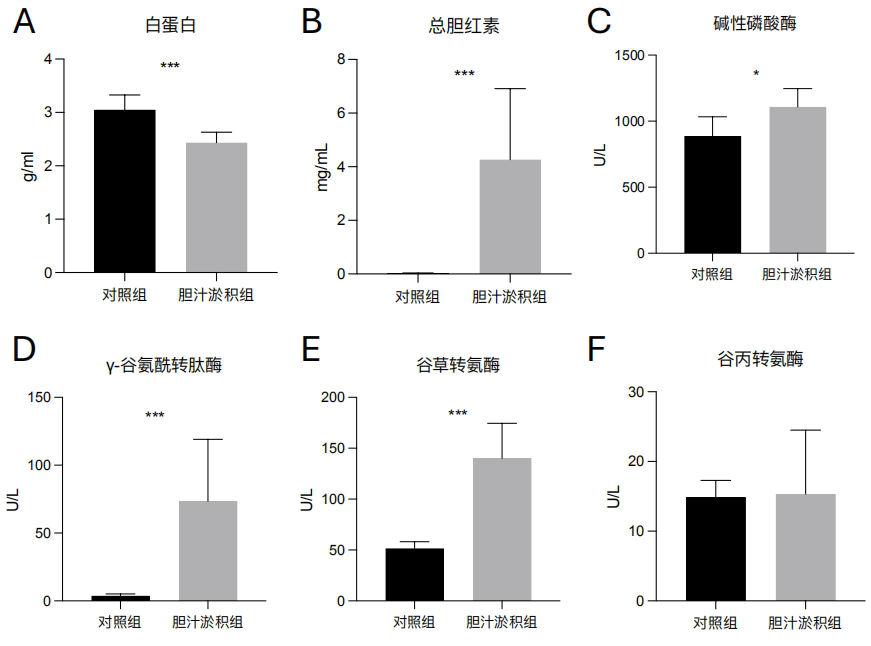
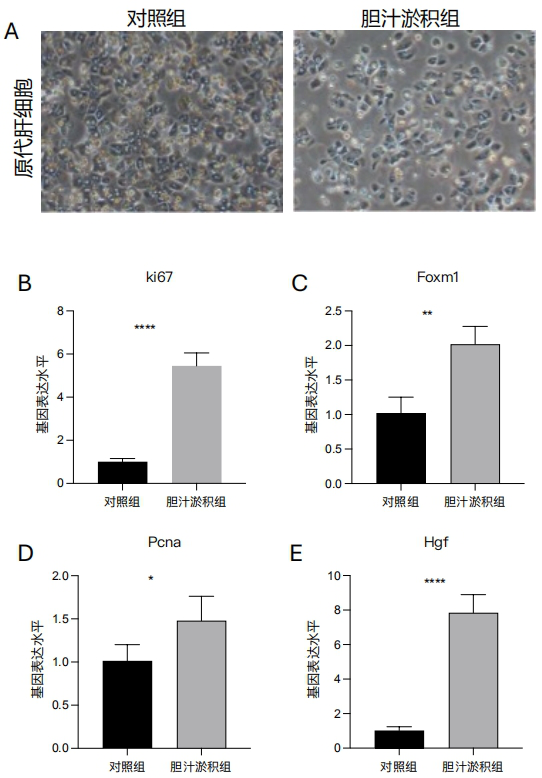
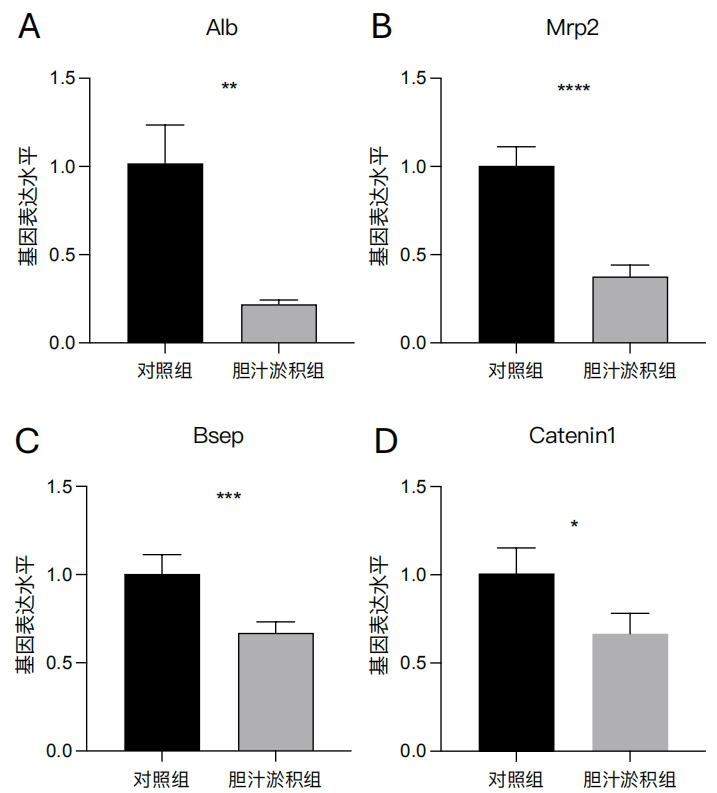
1、中华医学会肝病学分会.胆汁淤积性肝病管理指南(2021)[J].临床肝胆病杂志,2022,38(1):62-69.中华医学会肝病学分会.胆汁淤积性肝病管理指南(2021)[J].临床肝胆病杂志,2022,38(1):62-69.
2、沈斐斐,陆伦根.胆汁淤积性肝病的病因[J].实用肝脏病杂志,2016,19(6):644-646.沈斐斐,陆伦根.胆汁淤积性肝病的病因[J].实用肝脏病杂志,2016,19(6):644-646.
3、SATO%E2%80%83K%EF%BC%8CMARZIONI%E2%80%83M%EF%BC%8CMENG%E2%80%83F%EF%BC%8Cet%E2%80%83al%EF%BC%8EDuctular%E2%80%83%0Areaction%E2%80%83in%E2%80%83liver%E2%80%83diseases%EF%BC%9APathological%E2%80%83mechanisms%E2%80%83%0Aand%E2%80%83translational%E2%80%83significances%EF%BC%BBJ%EF%BC%BD%EF%BC%8EHepatology%EF%BC%8C%0A2019%EF%BC%8C69%EF%BC%881%EF%BC%89%EF%BC%9A420-430%EF%BC%8ESATO%E2%80%83K%EF%BC%8CMARZIONI%E2%80%83M%EF%BC%8CMENG%E2%80%83F%EF%BC%8Cet%E2%80%83al%EF%BC%8EDuctular%E2%80%83%0Areaction%E2%80%83in%E2%80%83liver%E2%80%83diseases%EF%BC%9APathological%E2%80%83mechanisms%E2%80%83%0Aand%E2%80%83translational%E2%80%83significances%EF%BC%BBJ%EF%BC%BD%EF%BC%8EHepatology%EF%BC%8C%0A2019%EF%BC%8C69%EF%BC%881%EF%BC%89%EF%BC%9A420-430%EF%BC%8E
4、TERADA%E2%80%83M%EF%BC%8CHORISAWA%E2%80%83K%EF%BC%8CMIURA%E2%80%83S%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AKupffer%E2%80%83%20cells%E2%80%83induce%E2%80%83%20Notch-mediated%E2%80%83%20hepatocyte%E2%80%83%0Aconversion%E2%80%83in%E2%80%83a%E2%80%83common%E2%80%83mouse%E2%80%83model%E2%80%83of%E2%80%83intrahepatic%E2%80%83%0Acholangiocarcinoma%EF%BC%BBJ%EF%BC%BD%EF%BC%8ESci%E2%80%83Rep%EF%BC%8C2016%EF%BC%886%EF%BC%89%EF%BC%9A%0A34691%EF%BC%8ETERADA%E2%80%83M%EF%BC%8CHORISAWA%E2%80%83K%EF%BC%8CMIURA%E2%80%83S%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AKupffer%E2%80%83%20cells%E2%80%83induce%E2%80%83%20Notch-mediated%E2%80%83%20hepatocyte%E2%80%83%0Aconversion%E2%80%83in%E2%80%83a%E2%80%83common%E2%80%83mouse%E2%80%83model%E2%80%83of%E2%80%83intrahepatic%E2%80%83%0Acholangiocarcinoma%EF%BC%BBJ%EF%BC%BD%EF%BC%8ESci%E2%80%83Rep%EF%BC%8C2016%EF%BC%886%EF%BC%89%EF%BC%9A%0A34691%EF%BC%8E
5、HU%E2%80%83S%EF%BC%8CMOLINA%E2%80%83L%EF%BC%8CTAO%E2%80%83J%EF%BC%8Cet%E2%80%83al%EF%BC%8ENOTCH-YAP1%2F%0ATEAD-DNMT1%E2%80%83axis%E2%80%83drives%E2%80%83hepatocyte%E2%80%83%20reprogramming%E2%80%83%0Ainto%E2%80%83intrahepatic%E2%80%83cholangiocarcinoma%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AGastroenterology%EF%BC%8C2022%EF%BC%8C163%EF%BC%882%EF%BC%89%EF%BC%9A449-465%EF%BC%8EHU%E2%80%83S%EF%BC%8CMOLINA%E2%80%83L%EF%BC%8CTAO%E2%80%83J%EF%BC%8Cet%E2%80%83al%EF%BC%8ENOTCH-YAP1%2F%0ATEAD-DNMT1%E2%80%83axis%E2%80%83drives%E2%80%83hepatocyte%E2%80%83%20reprogramming%E2%80%83%0Ainto%E2%80%83intrahepatic%E2%80%83cholangiocarcinoma%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AGastroenterology%EF%BC%8C2022%EF%BC%8C163%EF%BC%882%EF%BC%89%EF%BC%9A449-465%EF%BC%8E
6、SEKIYA%E2%80%83S%EF%BC%8CSUZUKI%E2%80%83A%EF%BC%8EHepatocytes%EF%BC%8Crather%E2%80%83than%E2%80%83%0Acholangiocytes%EF%BC%8Ccan%E2%80%83be%E2%80%83the%E2%80%83major%E2%80%83source%E2%80%83of%E2%80%83primitive%E2%80%83%0Aductules%E2%80%83in%E2%80%83the%E2%80%83chronically%E2%80%83injured%E2%80%83mouse%E2%80%83liver%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AAm%E2%80%83J%E2%80%83Pathol%EF%BC%8C2014%EF%BC%8C184%EF%BC%885%EF%BC%89%EF%BC%9A1468-1478%EF%BC%8ESEKIYA%E2%80%83S%EF%BC%8CSUZUKI%E2%80%83A%EF%BC%8EHepatocytes%EF%BC%8Crather%E2%80%83than%E2%80%83%0Acholangiocytes%EF%BC%8Ccan%E2%80%83be%E2%80%83the%E2%80%83major%E2%80%83source%E2%80%83of%E2%80%83primitive%E2%80%83%0Aductules%E2%80%83in%E2%80%83the%E2%80%83chronically%E2%80%83injured%E2%80%83mouse%E2%80%83liver%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AAm%E2%80%83J%E2%80%83Pathol%EF%BC%8C2014%EF%BC%8C184%EF%BC%885%EF%BC%89%EF%BC%9A1468-1478%EF%BC%8E
7、SUZUKI%E2%80%83A%EF%BC%8EEvidence%E2%80%83%20of%E2%80%83%20cell-fate%E2%80%83%20conversion%E2%80%83from%E2%80%83%0Ahepatocytes%E2%80%83to%E2%80%83cholangiocytes%E2%80%83in%E2%80%83the%E2%80%83injured%E2%80%83liver%EF%BC%9AIn%02vivo%E2%80%83genetic%E2%80%83lineage-tracing%E2%80%83approaches%EF%BC%BBJ%EF%BC%BD%EF%BC%8ECurr%E2%80%83%0AOpin%E2%80%83Gastroenterol%EF%BC%8C2015%EF%BC%8C31%EF%BC%883%EF%BC%89%EF%BC%9A247-251%EF%BC%8ESUZUKI%E2%80%83A%EF%BC%8EEvidence%E2%80%83%20of%E2%80%83%20cell-fate%E2%80%83%20conversion%E2%80%83from%E2%80%83%0Ahepatocytes%E2%80%83to%E2%80%83cholangiocytes%E2%80%83in%E2%80%83the%E2%80%83injured%E2%80%83liver%EF%BC%9AIn%02vivo%E2%80%83genetic%E2%80%83lineage-tracing%E2%80%83approaches%EF%BC%BBJ%EF%BC%BD%EF%BC%8ECurr%E2%80%83%0AOpin%E2%80%83Gastroenterol%EF%BC%8C2015%EF%BC%8C31%EF%BC%883%EF%BC%89%EF%BC%9A247-251%EF%BC%8E
8、HUANG%E2%80%83D%EF%BC%8CTANG%E2%80%83L%EF%BC%8CLI%E2%80%83T%EF%BC%8Cet%E2%80%83al%EF%BC%8EReversine%E2%80%83%0Aattenuates%E2%80%83cholestatic%E2%80%83ductular%E2%80%83reaction%E2%80%83in%E2%80%83rats%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AFEBS%E2%80%83Open%E2%80%83Bio%EF%BC%8C2023%EF%BC%8C13%EF%BC%885%EF%BC%89%EF%BC%9A898-911%EF%BC%8EHUANG%E2%80%83D%EF%BC%8CTANG%E2%80%83L%EF%BC%8CLI%E2%80%83T%EF%BC%8Cet%E2%80%83al%EF%BC%8EReversine%E2%80%83%0Aattenuates%E2%80%83cholestatic%E2%80%83ductular%E2%80%83reaction%E2%80%83in%E2%80%83rats%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AFEBS%E2%80%83Open%E2%80%83Bio%EF%BC%8C2023%EF%BC%8C13%EF%BC%885%EF%BC%89%EF%BC%9A898-911%EF%BC%8E
9、ZHANG%E2%80%83S%EF%BC%8CHUANG%E2%80%83D%EF%BC%8CWENG%E2%80%83J%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ANeutralization%E2%80%83of%E2%80%83interleukin%E2%80%9017%E2%80%83attenuates%E2%80%83cholestatic%E2%80%83%0Aliver%E2%80%83fibrosis%E2%80%83in%E2%80%83mice%EF%BC%BBJ%EF%BC%BD%EF%BC%8EScand%E2%80%83J%E2%80%83Immunol%EF%BC%8C%0A2016%EF%BC%8C83%EF%BC%882%EF%BC%89%EF%BC%9A102-108%EF%BC%8EZHANG%E2%80%83S%EF%BC%8CHUANG%E2%80%83D%EF%BC%8CWENG%E2%80%83J%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ANeutralization%E2%80%83of%E2%80%83interleukin%E2%80%9017%E2%80%83attenuates%E2%80%83cholestatic%E2%80%83%0Aliver%E2%80%83fibrosis%E2%80%83in%E2%80%83mice%EF%BC%BBJ%EF%BC%BD%EF%BC%8EScand%E2%80%83J%E2%80%83Immunol%EF%BC%8C%0A2016%EF%BC%8C83%EF%BC%882%EF%BC%89%EF%BC%9A102-108%EF%BC%8E
10、HUANG%E2%80%83Y%EF%BC%8CSAKAI%E2%80%83Y%EF%BC%8CHARA%E2%80%83T%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ABioengineering%E2%80%83of%E2%80%83a%E2%80%83CLiP%E2%80%90derived%E2%80%83tubular%E2%80%83biliary%E2%80%90%0Aduct%E2%80%90like%E2%80%83structure%E2%80%83for%E2%80%83bile%E2%80%83transport%E2%80%83in%E2%80%83vitro%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ABiotechnol%E2%80%83Bioeng%EF%BC%8C2021%EF%BC%8C118%EF%BC%887%EF%BC%89%EF%BC%9A2572-2584%EF%BC%8EHUANG%E2%80%83Y%EF%BC%8CSAKAI%E2%80%83Y%EF%BC%8CHARA%E2%80%83T%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ABioengineering%E2%80%83of%E2%80%83a%E2%80%83CLiP%E2%80%90derived%E2%80%83tubular%E2%80%83biliary%E2%80%90%0Aduct%E2%80%90like%E2%80%83structure%E2%80%83for%E2%80%83bile%E2%80%83transport%E2%80%83in%E2%80%83vitro%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ABiotechnol%E2%80%83Bioeng%EF%BC%8C2021%EF%BC%8C118%EF%BC%887%EF%BC%89%EF%BC%9A2572-2584%EF%BC%8E
11、HUANG%E2%80%83Y%EF%BC%8CMIYAMOTO%E2%80%83D%EF%BC%8CLI%E2%80%83P%E2%80%83L%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AChemical%E2%80%83conversion%E2%80%83of%E2%80%83aged%E2%80%83hepatocytes%E2%80%83into%E2%80%83bipotent%E2%80%83%0Aliver%E2%80%83progenitor%E2%80%83cells%EF%BC%BBJ%EF%BC%BD%EF%BC%8EHepatol%E2%80%83Res%EF%BC%8C2021%EF%BC%8C51%0A%EF%BC%883%EF%BC%89%EF%BC%9A323-335%EF%BC%8EHUANG%E2%80%83Y%EF%BC%8CMIYAMOTO%E2%80%83D%EF%BC%8CLI%E2%80%83P%E2%80%83L%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AChemical%E2%80%83conversion%E2%80%83of%E2%80%83aged%E2%80%83hepatocytes%E2%80%83into%E2%80%83bipotent%E2%80%83%0Aliver%E2%80%83progenitor%E2%80%83cells%EF%BC%BBJ%EF%BC%BD%EF%BC%8EHepatol%E2%80%83Res%EF%BC%8C2021%EF%BC%8C51%0A%EF%BC%883%EF%BC%89%EF%BC%9A323-335%EF%BC%8E
12、SCHMITTGEN%E2%80%83T%E2%80%83D%EF%BC%8CLIVAK%E2%80%83K%E2%80%83J%EF%BC%8EAnalyzing%E2%80%83real-time%E2%80%83%0APCR%E2%80%83data%E2%80%83by%E2%80%83the%E2%80%83comparative%E2%80%83C%EF%BC%88T%EF%BC%89method%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENat%E2%80%83%0AProtoc%EF%BC%8C2008%EF%BC%8C3%EF%BC%886%EF%BC%89%EF%BC%9A1101-1108%EF%BC%8ESCHMITTGEN%E2%80%83T%E2%80%83D%EF%BC%8CLIVAK%E2%80%83K%E2%80%83J%EF%BC%8EAnalyzing%E2%80%83real-time%E2%80%83%0APCR%E2%80%83data%E2%80%83by%E2%80%83the%E2%80%83comparative%E2%80%83C%EF%BC%88T%EF%BC%89method%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENat%E2%80%83%0AProtoc%EF%BC%8C2008%EF%BC%8C3%EF%BC%886%EF%BC%89%EF%BC%9A1101-1108%EF%BC%8E
13、FUKASAWA%E2%80%83H%EF%BC%8CHASHIMOTO%E2%80%83R%EF%BC%8CHAGIHARA%E2%80%83%0AK%EF%BC%8Cet%E2%80%83al%EF%BC%8EHistological%E2%80%83%20species%E2%80%83%20differences%E2%80%83%20among%E2%80%83%0Achickens%EF%BC%8Crats%EF%BC%8Cand%E2%80%83mice%E2%80%83in%E2%80%83experimental%E2%80%83cholestasis%E2%80%83%0Aby%E2%80%83bile%E2%80%83duct%E2%80%83ligation%EF%BC%BBJ%EF%BC%BD%EF%BC%8ERes%E2%80%83Vet%E2%80%83Sci%EF%BC%8C2024%0A%EF%BC%88176%EF%BC%89%EF%BC%9A105343%EF%BC%8EFUKASAWA%E2%80%83H%EF%BC%8CHASHIMOTO%E2%80%83R%EF%BC%8CHAGIHARA%E2%80%83%0AK%EF%BC%8Cet%E2%80%83al%EF%BC%8EHistological%E2%80%83%20species%E2%80%83%20differences%E2%80%83%20among%E2%80%83%0Achickens%EF%BC%8Crats%EF%BC%8Cand%E2%80%83mice%E2%80%83in%E2%80%83experimental%E2%80%83cholestasis%E2%80%83%0Aby%E2%80%83bile%E2%80%83duct%E2%80%83ligation%EF%BC%BBJ%EF%BC%BD%EF%BC%8ERes%E2%80%83Vet%E2%80%83Sci%EF%BC%8C2024%0A%EF%BC%88176%EF%BC%89%EF%BC%9A105343%EF%BC%8E
14、TANG%E2%80%83G%EF%BC%8CNIERATH%E2%80%83W%E2%80%83F%EF%BC%8CLEITNER%E2%80%83E%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AComparing%E2%80%83%20animal%E2%80%83%20well-being%E2%80%83%20between%E2%80%83%20bile%E2%80%83%20duct%E2%80%83%0Aligation%E2%80%83models%EF%BC%BBJ%EF%BC%BD%EF%BC%8EPLoS%E2%80%83One%EF%BC%8C2024%EF%BC%8C19%EF%BC%887%EF%BC%89%EF%BC%9A%0Ae0303786%EF%BC%8ETANG%E2%80%83G%EF%BC%8CNIERATH%E2%80%83W%E2%80%83F%EF%BC%8CLEITNER%E2%80%83E%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AComparing%E2%80%83%20animal%E2%80%83%20well-being%E2%80%83%20between%E2%80%83%20bile%E2%80%83%20duct%E2%80%83%0Aligation%E2%80%83models%EF%BC%BBJ%EF%BC%BD%EF%BC%8EPLoS%E2%80%83One%EF%BC%8C2024%EF%BC%8C19%EF%BC%887%EF%BC%89%EF%BC%9A%0Ae0303786%EF%BC%8E
15、%E2%80%83%20LADERIAN%E2%80%83A%EF%BC%8CGHASEMI%E2%80%83M%EF%BC%8CMORTAZAVI%E2%80%83P%EF%BC%8C%0Aet%E2%80%83al%EF%BC%8EHepatoprotective%E2%80%83effect%E2%80%83of%E2%80%83astaxanthin%E2%80%83against%E2%80%83%0Acholestasis%E2%80%83liver%E2%80%83fibrosis%E2%80%83induced%E2%80%83by%E2%80%83bile%E2%80%83duct%E2%80%83ligation%E2%80%83%0Ain%E2%80%83adult%E2%80%83Wistar%E2%80%83rats%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Biochem%E2%80%83Mol%E2%80%83Toxicol%EF%BC%8C%0A2024%EF%BC%8C38%EF%BC%888%EF%BC%89%EF%BC%9Ae23788%EF%BC%8E%E2%80%83%20LADERIAN%E2%80%83A%EF%BC%8CGHASEMI%E2%80%83M%EF%BC%8CMORTAZAVI%E2%80%83P%EF%BC%8C%0Aet%E2%80%83al%EF%BC%8EHepatoprotective%E2%80%83effect%E2%80%83of%E2%80%83astaxanthin%E2%80%83against%E2%80%83%0Acholestasis%E2%80%83liver%E2%80%83fibrosis%E2%80%83induced%E2%80%83by%E2%80%83bile%E2%80%83duct%E2%80%83ligation%E2%80%83%0Ain%E2%80%83adult%E2%80%83Wistar%E2%80%83rats%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Biochem%E2%80%83Mol%E2%80%83Toxicol%EF%BC%8C%0A2024%EF%BC%8C38%EF%BC%888%EF%BC%89%EF%BC%9Ae23788%EF%BC%8E
16、GRIBBEN%E2%80%83C%EF%BC%8CGALANAKIS%E2%80%83V%EF%BC%8CCALDERWOOD%E2%80%83A%EF%BC%8C%0Aet%E2%80%83al%EF%BC%8EAcquisition%E2%80%83%20of%E2%80%83%20epithelial%E2%80%83%20plasticity%E2%80%83in%E2%80%83%20human%E2%80%83%0Achronic%E2%80%83liver%E2%80%83disease%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENature%EF%BC%8C2024%EF%BC%8C630%0A%EF%BC%888015%EF%BC%89%EF%BC%9A166-173%EF%BC%8EGRIBBEN%E2%80%83C%EF%BC%8CGALANAKIS%E2%80%83V%EF%BC%8CCALDERWOOD%E2%80%83A%EF%BC%8C%0Aet%E2%80%83al%EF%BC%8EAcquisition%E2%80%83%20of%E2%80%83%20epithelial%E2%80%83%20plasticity%E2%80%83in%E2%80%83%20human%E2%80%83%0Achronic%E2%80%83liver%E2%80%83disease%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENature%EF%BC%8C2024%EF%BC%8C630%0A%EF%BC%888015%EF%BC%89%EF%BC%9A166-173%EF%BC%8E
17、TARLOW%E2%80%83B%E2%80%83D%EF%BC%8CPELZ%E2%80%83C%EF%BC%8CNAUGLER%E2%80%83W%E2%80%83E%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ABipotential%E2%80%83%20adult%E2%80%83liver%E2%80%83%20progenitors%E2%80%83%20are%E2%80%83%20derived%E2%80%83from%E2%80%83%0Achronically%E2%80%83injured%E2%80%83mature%E2%80%83hepatocytes%EF%BC%BBJ%EF%BC%BD%EF%BC%8ECell%E2%80%83%0AStem%E2%80%83Cell%EF%BC%8C2014%EF%BC%8C15%EF%BC%885%EF%BC%89%EF%BC%9A605-618%EF%BC%8ETARLOW%E2%80%83B%E2%80%83D%EF%BC%8CPELZ%E2%80%83C%EF%BC%8CNAUGLER%E2%80%83W%E2%80%83E%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ABipotential%E2%80%83%20adult%E2%80%83liver%E2%80%83%20progenitors%E2%80%83%20are%E2%80%83%20derived%E2%80%83from%E2%80%83%0Achronically%E2%80%83injured%E2%80%83mature%E2%80%83hepatocytes%EF%BC%BBJ%EF%BC%BD%EF%BC%8ECell%E2%80%83%0AStem%E2%80%83Cell%EF%BC%8C2014%EF%BC%8C15%EF%BC%885%EF%BC%89%EF%BC%9A605-618%EF%BC%8E
18、YANGER%E2%80%83K%EF%BC%8CZONG%E2%80%83Y%EF%BC%8CMAGGS%E2%80%83L%E2%80%83R%EF%BC%8Cet%E2%80%83al%EF%BC%8ERobust%E2%80%83%0Acellular%E2%80%83%20reprogramming%E2%80%83occurs%E2%80%83%20spontaneously%E2%80%83%20during%E2%80%83%0Aliver%E2%80%83regeneration%EF%BC%BBJ%EF%BC%BD%EF%BC%8EGenes%E2%80%83Dev%EF%BC%8C2013%EF%BC%8C27%0A%EF%BC%887%EF%BC%89%EF%BC%9A719-724%EF%BC%8EYANGER%E2%80%83K%EF%BC%8CZONG%E2%80%83Y%EF%BC%8CMAGGS%E2%80%83L%E2%80%83R%EF%BC%8Cet%E2%80%83al%EF%BC%8ERobust%E2%80%83%0Acellular%E2%80%83%20reprogramming%E2%80%83occurs%E2%80%83%20spontaneously%E2%80%83%20during%E2%80%83%0Aliver%E2%80%83regeneration%EF%BC%BBJ%EF%BC%BD%EF%BC%8EGenes%E2%80%83Dev%EF%BC%8C2013%EF%BC%8C27%0A%EF%BC%887%EF%BC%89%EF%BC%9A719-724%EF%BC%8E
19、AGUILAR-BRAVO%E2%80%83B%EF%BC%8CARINO%E2%80%83S%EF%BC%8CBLAYA%E2%80%83D%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AHepatocyte%E2%80%83dedifferentiation%E2%80%83profiling%E2%80%83in%E2%80%83alcohol%02related%E2%80%83liver%E2%80%83disease%E2%80%83identifies%E2%80%83CXCR4%E2%80%83as%E2%80%83a%E2%80%83driver%E2%80%83of%E2%80%83cell%E2%80%83%0Areprogramming%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Hepatol%EF%BC%8C2023%EF%BC%8C79%EF%BC%883%EF%BC%89%EF%BC%9A%0A728-740%EF%BC%8EAGUILAR-BRAVO%E2%80%83B%EF%BC%8CARINO%E2%80%83S%EF%BC%8CBLAYA%E2%80%83D%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AHepatocyte%E2%80%83dedifferentiation%E2%80%83profiling%E2%80%83in%E2%80%83alcohol%02related%E2%80%83liver%E2%80%83disease%E2%80%83identifies%E2%80%83CXCR4%E2%80%83as%E2%80%83a%E2%80%83driver%E2%80%83of%E2%80%83cell%E2%80%83%0Areprogramming%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Hepatol%EF%BC%8C2023%EF%BC%8C79%EF%BC%883%EF%BC%89%EF%BC%9A%0A728-740%EF%BC%8E
20、RAVEN%E2%80%83A%EF%BC%8CLU%E2%80%83W%E2%80%83Y%EF%BC%8CMAN%E2%80%83T%E2%80%83Y%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ACholangiocytes%E2%80%83act%E2%80%83as%E2%80%83facultative%E2%80%83liver%E2%80%83stem%E2%80%83cells%E2%80%83during%E2%80%83%0Aimpaired%E2%80%83hepatocyte%E2%80%83regeneration%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENature%EF%BC%8C%0A2017%EF%BC%8C547%EF%BC%887663%EF%BC%89%EF%BC%9A350-354%EF%BC%8ERAVEN%E2%80%83A%EF%BC%8CLU%E2%80%83W%E2%80%83Y%EF%BC%8CMAN%E2%80%83T%E2%80%83Y%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0ACholangiocytes%E2%80%83act%E2%80%83as%E2%80%83facultative%E2%80%83liver%E2%80%83stem%E2%80%83cells%E2%80%83during%E2%80%83%0Aimpaired%E2%80%83hepatocyte%E2%80%83regeneration%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENature%EF%BC%8C%0A2017%EF%BC%8C547%EF%BC%887663%EF%BC%89%EF%BC%9A350-354%EF%BC%8E
21、HE%E2%80%83J%EF%BC%8CLU%E2%80%83H%EF%BC%8CZOU%E2%80%83Q%EF%BC%8Cet%E2%80%83al%EF%BC%8ERegeneration%E2%80%83%20of%E2%80%83%0Aliver%E2%80%83%20after%E2%80%83%20extreme%E2%80%83%20hepatocyte%E2%80%83loss%E2%80%83%20occurs%E2%80%83%20mainly%E2%80%83%0Avia%E2%80%83biliary%E2%80%83transdifferentiation%E2%80%83in%E2%80%83zebrafish%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AGastroenterology%EF%BC%8C2014%EF%BC%8C146%EF%BC%883%EF%BC%89%EF%BC%9A789-800%EF%BC%8Ee8%EF%BC%8EHE%E2%80%83J%EF%BC%8CLU%E2%80%83H%EF%BC%8CZOU%E2%80%83Q%EF%BC%8Cet%E2%80%83al%EF%BC%8ERegeneration%E2%80%83%20of%E2%80%83%0Aliver%E2%80%83%20after%E2%80%83%20extreme%E2%80%83%20hepatocyte%E2%80%83loss%E2%80%83%20occurs%E2%80%83%20mainly%E2%80%83%0Avia%E2%80%83biliary%E2%80%83transdifferentiation%E2%80%83in%E2%80%83zebrafish%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AGastroenterology%EF%BC%8C2014%EF%BC%8C146%EF%BC%883%EF%BC%89%EF%BC%9A789-800%EF%BC%8Ee8%EF%BC%8E
22、%E2%80%83%20MICHALOPOULOS%E2%80%83G%E2%80%83K%EF%BC%8CBARUA%E2%80%83L%EF%BC%8CBOWEN%E2%80%83W%E2%80%83C%EF%BC%8E%0ATransdifferentiation%E2%80%83of%E2%80%83rat%E2%80%83hepatocytes%E2%80%83into%E2%80%83biliary%E2%80%83cells%E2%80%83%0Aafter%E2%80%83bile%E2%80%83duct%E2%80%83ligation%E2%80%83and%E2%80%83toxic%E2%80%83biliary%E2%80%83injury%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AHepatology%EF%BC%8C2005%EF%BC%8C41%EF%BC%883%EF%BC%89%EF%BC%9A535-544%EF%BC%8E%E2%80%83%20MICHALOPOULOS%E2%80%83G%E2%80%83K%EF%BC%8CBARUA%E2%80%83L%EF%BC%8CBOWEN%E2%80%83W%E2%80%83C%EF%BC%8E%0ATransdifferentiation%E2%80%83of%E2%80%83rat%E2%80%83hepatocytes%E2%80%83into%E2%80%83biliary%E2%80%83cells%E2%80%83%0Aafter%E2%80%83bile%E2%80%83duct%E2%80%83ligation%E2%80%83and%E2%80%83toxic%E2%80%83biliary%E2%80%83injury%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0AHepatology%EF%BC%8C2005%EF%BC%8C41%EF%BC%883%EF%BC%89%EF%BC%9A535-544%EF%BC%8E
23、YOVCHEV%E2%80%83M%E2%80%83I%EF%BC%8CLOCKER%E2%80%83J%EF%BC%8COERTEL%E2%80%83M%EF%BC%8EBiliary%E2%80%83%0Afibrosis%E2%80%83%20drives%E2%80%83%20liver%E2%80%83%20repopulation%E2%80%83%20and%E2%80%83%20phenotype%E2%80%83%0Atransition%E2%80%83of%E2%80%83transplanted%E2%80%83hepatocytes%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Hepatol%EF%BC%8C2016%EF%BC%8C64%EF%BC%886%EF%BC%89%EF%BC%9A1348-1357%EF%BC%8EYOVCHEV%E2%80%83M%E2%80%83I%EF%BC%8CLOCKER%E2%80%83J%EF%BC%8COERTEL%E2%80%83M%EF%BC%8EBiliary%E2%80%83%0Afibrosis%E2%80%83%20drives%E2%80%83%20liver%E2%80%83%20repopulation%E2%80%83%20and%E2%80%83%20phenotype%E2%80%83%0Atransition%E2%80%83of%E2%80%83transplanted%E2%80%83hepatocytes%EF%BC%BBJ%EF%BC%BD%EF%BC%8EJ%E2%80%83Hepatol%EF%BC%8C2016%EF%BC%8C64%EF%BC%886%EF%BC%89%EF%BC%9A1348-1357%EF%BC%8E
24、%E2%80%83%20FONT-BURGADA%E2%80%83J%EF%BC%8CSHALAPOUR%E2%80%83S%EF%BC%8CRAMASWAMY%E2%80%83%0AS%EF%BC%8Cet%E2%80%83al%EF%BC%8EHybrid%E2%80%83%20periportal%E2%80%83%20hepatocytes%E2%80%83%20regenerate%E2%80%83%0Athe%E2%80%83injured%E2%80%83liver%E2%80%83without%E2%80%83giving%E2%80%83rise%E2%80%83to%E2%80%83cancer%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ACell%EF%BC%8C2015%EF%BC%8C162%EF%BC%884%EF%BC%89%EF%BC%9A766-779%EF%BC%8E%E2%80%83%20FONT-BURGADA%E2%80%83J%EF%BC%8CSHALAPOUR%E2%80%83S%EF%BC%8CRAMASWAMY%E2%80%83%0AS%EF%BC%8Cet%E2%80%83al%EF%BC%8EHybrid%E2%80%83%20periportal%E2%80%83%20hepatocytes%E2%80%83%20regenerate%E2%80%83%0Athe%E2%80%83injured%E2%80%83liver%E2%80%83without%E2%80%83giving%E2%80%83rise%E2%80%83to%E2%80%83cancer%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ACell%EF%BC%8C2015%EF%BC%8C162%EF%BC%884%EF%BC%89%EF%BC%9A766-779%EF%BC%8E
25、邱冰峰,都金星,申定珠,等.肝细胞向胆管上皮细胞转分化在继发性胆汁淤积性大鼠肝纤维化形成中的作用及黄芪汤组分的干预效应[J].中国中西医结合杂志,2010,30(5):513-518.邱冰峰,都金星,申定珠,等.肝细胞向胆管上皮细胞转分化在继发性胆汁淤积性大鼠肝纤维化形成中的作用及黄芪汤组分的干预效应[J].中国中西医结合杂志,2010,30(5):513-518.
26、%E2%80%83%20TANG%E2%80%83D%EF%BC%8CCHEN%E2%80%83Y%EF%BC%8CFU%E2%80%83G%E2%80%83B%EF%BC%8Cet%E2%80%83al%EF%BC%8EEpCAM%E2%80%83inhibits%E2%80%83%0Adifferentiation%E2%80%83%20of%E2%80%83%20human%E2%80%83liver%E2%80%83%20progenitor%E2%80%83%20cells%E2%80%83into%E2%80%83%0Ahepatocytes%E2%80%83in%E2%80%83vitro%E2%80%83by%E2%80%83activating%E2%80%83Notch1%E2%80%83signaling%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ABiochem%E2%80%83Biophys%E2%80%83Res%E2%80%83Commun%EF%BC%8C2020%EF%BC%8CS0006-291X%0A%EF%BC%8820%EF%BC%8930309-0%EF%BC%8E%E2%80%83%20TANG%E2%80%83D%EF%BC%8CCHEN%E2%80%83Y%EF%BC%8CFU%E2%80%83G%E2%80%83B%EF%BC%8Cet%E2%80%83al%EF%BC%8EEpCAM%E2%80%83inhibits%E2%80%83%0Adifferentiation%E2%80%83%20of%E2%80%83%20human%E2%80%83liver%E2%80%83%20progenitor%E2%80%83%20cells%E2%80%83into%E2%80%83%0Ahepatocytes%E2%80%83in%E2%80%83vitro%E2%80%83by%E2%80%83activating%E2%80%83Notch1%E2%80%83signaling%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ABiochem%E2%80%83Biophys%E2%80%83Res%E2%80%83Commun%EF%BC%8C2020%EF%BC%8CS0006-291X%0A%EF%BC%8820%EF%BC%8930309-0%EF%BC%8E
27、SHANG%E2%80%83T%EF%BC%8CJIANG%E2%80%83T%EF%BC%8CCUI%E2%80%83X%EF%BC%8Cet%E2%80%83al%EF%BC%8EDive%20rse%E2%80%83%0Afunctions%E2%80%83of%E2%80%83SOX9%E2%80%83in%E2%80%83liver%E2%80%83development%E2%80%83and%E2%80%83homeostasis%E2%80%83%0Aand%E2%80%83hepatobiliary%E2%80%83diseases%EF%BC%BBJ%EF%BC%BD%EF%BC%8EGenes%E2%80%83Dis%EF%BC%8C2024%EF%BC%8C%0A11%EF%BC%884%EF%BC%89%EF%BC%9A100996%EF%BC%8ESHANG%E2%80%83T%EF%BC%8CJIANG%E2%80%83T%EF%BC%8CCUI%E2%80%83X%EF%BC%8Cet%E2%80%83al%EF%BC%8EDive%20rse%E2%80%83%0Afunctions%E2%80%83of%E2%80%83SOX9%E2%80%83in%E2%80%83liver%E2%80%83development%E2%80%83and%E2%80%83homeostasis%E2%80%83%0Aand%E2%80%83hepatobiliary%E2%80%83diseases%EF%BC%BBJ%EF%BC%BD%EF%BC%8EGenes%E2%80%83Dis%EF%BC%8C2024%EF%BC%8C%0A11%EF%BC%884%EF%BC%89%EF%BC%9A100996%EF%BC%8E
28、LEE%E2%80%83S%E2%80%83H%20%EF%BC%8C%20SO%E2%80%83J%20%EF%BC%8C%20SHIN%E2%80%83D%20%EF%BC%8E%20Hepatocyte%02t%20o%20-%20c%20h%20ol%20a%20n%20gi%20o%20c%20yt%20e%E2%80%83%20c%20o%20n%20v%20e%20r%20si%20o%20n%E2%80%83%20o%20c%20c%20u%20r%20s%E2%80%83%20t%20h%20r%20o%20u%20g%20h%E2%80%83%0Atransdifferentiation%E2%80%83independently%E2%80%83of%E2%80%83%20proliferation%E2%80%83in%E2%80%83%0Azebrafish%EF%BC%BBJ%EF%BC%BD%EF%BC%8EHepatology%EF%BC%8C2023%EF%BC%8C77%EF%BC%884%EF%BC%89%EF%BC%9A%0A1198-1210%EF%BC%8ELEE%E2%80%83S%E2%80%83H%20%EF%BC%8C%20SO%E2%80%83J%20%EF%BC%8C%20SHIN%E2%80%83D%20%EF%BC%8E%20Hepatocyte%02t%20o%20-%20c%20h%20ol%20a%20n%20gi%20o%20c%20yt%20e%E2%80%83%20c%20o%20n%20v%20e%20r%20si%20o%20n%E2%80%83%20o%20c%20c%20u%20r%20s%E2%80%83%20t%20h%20r%20o%20u%20g%20h%E2%80%83%0Atransdifferentiation%E2%80%83independently%E2%80%83of%E2%80%83%20proliferation%E2%80%83in%E2%80%83%0Azebrafish%EF%BC%BBJ%EF%BC%BD%EF%BC%8EHepatology%EF%BC%8C2023%EF%BC%8C77%EF%BC%884%EF%BC%89%EF%BC%9A%0A1198-1210%EF%BC%8E
29、LAN%E2%80%83T%EF%BC%8CTAI%E2%80%83Y%EF%BC%8CZHAO%E2%80%83C%EF%BC%8Cet%E2%80%83al%EF%BC%8EAt%20y%20pi%20c%20al%E2%80%83%0Acholangiocytes%E2%80%83derived%E2%80%83from%E2%80%83hepatocyte-cholangiocyte%E2%80%83%0Atransdifferentiation%E2%80%83mediated%E2%80%83by%E2%80%83COX-2%EF%BC%9AA%E2%80%83%20kind%E2%80%83of%E2%80%83%0Amisguided%E2%80%83liver%E2%80%83regeneration%EF%BC%BBJ%EF%BC%BD%EF%BC%8EInflamm%E2%80%83Regen%EF%BC%8C%0A2023%EF%BC%8C43%EF%BC%881%EF%BC%89%EF%BC%9A37%EF%BC%8ELAN%E2%80%83T%EF%BC%8CTAI%E2%80%83Y%EF%BC%8CZHAO%E2%80%83C%EF%BC%8Cet%E2%80%83al%EF%BC%8EAt%20y%20pi%20c%20al%E2%80%83%0Acholangiocytes%E2%80%83derived%E2%80%83from%E2%80%83hepatocyte-cholangiocyte%E2%80%83%0Atransdifferentiation%E2%80%83mediated%E2%80%83by%E2%80%83COX-2%EF%BC%9AA%E2%80%83%20kind%E2%80%83of%E2%80%83%0Amisguided%E2%80%83liver%E2%80%83regeneration%EF%BC%BBJ%EF%BC%BD%EF%BC%8EInflamm%E2%80%83Regen%EF%BC%8C%0A2023%EF%BC%8C43%EF%BC%881%EF%BC%89%EF%BC%9A37%EF%BC%8E
30、CHEN%E2%80%83Y%EF%BC%8CYAN%E2%80%83Y%EF%BC%8CLI%E2%80%83Y%EF%BC%8Cet%E2%80%83al%EF%BC%8EDeletion%E2%80%83%20of%E2%80%83%0ATgm2%E2%80%83suppresses%E2%80%83BMP-mediated%E2%80%83hepatocyte-to%02cholangiocyte%E2%80%83metaplasia%E2%80%83in%E2%80%83ductular%E2%80%83reaction%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ACell%E2%80%83Prolif%EF%BC%8C2024%EF%BC%8C57%EF%BC%8810%EF%BC%89%EF%BC%9Ae13646%EF%BC%8ECHEN%E2%80%83Y%EF%BC%8CYAN%E2%80%83Y%EF%BC%8CLI%E2%80%83Y%EF%BC%8Cet%E2%80%83al%EF%BC%8EDeletion%E2%80%83%20of%E2%80%83%0ATgm2%E2%80%83suppresses%E2%80%83BMP-mediated%E2%80%83hepatocyte-to%02cholangiocyte%E2%80%83metaplasia%E2%80%83in%E2%80%83ductular%E2%80%83reaction%EF%BC%BBJ%EF%BC%BD%EF%BC%8E%0ACell%E2%80%83Prolif%EF%BC%8C2024%EF%BC%8C57%EF%BC%8810%EF%BC%89%EF%BC%9Ae13646%EF%BC%8E
31、XU%E2%80%83J%EF%BC%8CKAUSALYA%E2%80%83P%E2%80%83J%EF%BC%8CONG%E2%80%83A%E2%80%83G%E2%80%83M%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AZO-2%2FTjp2%E2%80%83suppresses%E2%80%83Yap%E2%80%83and%E2%80%83Wwtr1%2FTaz-mediated%E2%80%83%0Ahepatocyte%E2%80%83to%E2%80%83%20cholangiocyte%E2%80%83transdifferentiation%E2%80%83in%E2%80%83%0Athe%E2%80%83mouse%E2%80%83liver%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENPJ%E2%80%83Regen%E2%80%83Med%EF%BC%8C2022%EF%BC%8C7%0A%EF%BC%881%EF%BC%89%EF%BC%9A55%EF%BC%8EXU%E2%80%83J%EF%BC%8CKAUSALYA%E2%80%83P%E2%80%83J%EF%BC%8CONG%E2%80%83A%E2%80%83G%E2%80%83M%EF%BC%8Cet%E2%80%83al%EF%BC%8E%0AZO-2%2FTjp2%E2%80%83suppresses%E2%80%83Yap%E2%80%83and%E2%80%83Wwtr1%2FTaz-mediated%E2%80%83%0Ahepatocyte%E2%80%83to%E2%80%83%20cholangiocyte%E2%80%83transdifferentiation%E2%80%83in%E2%80%83%0Athe%E2%80%83mouse%E2%80%83liver%EF%BC%BBJ%EF%BC%BD%EF%BC%8ENPJ%E2%80%83Regen%E2%80%83Med%EF%BC%8C2022%EF%BC%8C7%0A%EF%BC%881%EF%BC%89%EF%BC%9A55%EF%BC%8E
32、张启迪,陆伦根.肝细胞转分化是肝内胆汁淤积治疗的新方向[J].肝脏,2019,24(6):611.张启迪,陆伦根.肝细胞转分化是肝内胆汁淤积治疗的新方向[J].肝脏,2019,24(6):611.